Всем известно, что бурые медведи чрезвычайно опасны, и поэтому если хочется отдохнуть в местах их обитания, не стоит терять бдительность за прочтением книги...
Косолапые и неуклюжие медведи в цирке всегда поражали зрителей своим послушанием и умением выполнять трюки, не хуже других цирковых животных. Несмотря на свою...
Всем известная большая панда, имеет очень красивую родственницу – малую панду. Это животное имеет схожесть по внешнему виду с лисицей, у нее огненно-рыжий...
Само слово медведь, нередко ассоциируется с образом человека — могущественного, сильного, выносливого, способного делать такие вещи, которые неподвластны обычному народу, также помимо силы...
Черного медведя еще называют барибал. Это обыкновенный медведь Северной Америки, обитает на территории от Аляски до Канады и от побережья Тихого океана до...
Всем с детства известен добрый и музыкальный мультфильм про белого медвежонка Умку. Маленький белоснежный комок с черным носом никого не оставляет равнодушным. Увидеть...
Гималайский медведь – редкий подвид Азиатского черного медведя. Среда обитания гималайских черных медведей распространена на территории, простирающейся через Гималаи в Пакистан. Их наибольшее...
Во все времена охота на диких зверей занимала важное место в жизни людей. Постепенно из занятия, необходимого для получения пищи, охота превратилась в...
Бируанги или малайские медведи – самые маленькие представители среди медвежьих. В народе таких медведей называют медведями-собаками за определенной сходство с псовыми. Обитает бируанг...
У посетителей зоопарков самый живой интерес пробуждают человекообразные обезьяны, обитатели тропиков – слоны, жирафы, носороги, и крупные наземные хищники – медведи. Трехвековой опыт...
Популяция гималайских медведей в природе имеет довольно низкую численность с существенным риском полного исчезновения. Причинами сокращения их численности стала постоянная вырубка деревьев и...
Во всех широтах, где встречаются медведи, их сила, выносливость и ум являются предметом поклонения и особого восхищения у местных жителей. Изнеженные горожане, которые...
Местом обитания белогрудых медведей являются холмовые и горные леса, которые распростёрлись от Ирана, через Афганистан, а также Гималаи – от Кореи и Японии....
Белые медведи или по-другому полярные медведи, удивительно находчивые и умные млекопитающие, особенно когда добывают еду для себя и своих детей. Эти хищники самые...
Большая панда – это красивое и редкое животное с чёрно-белым окрасом. По своему телосложению она похожа на медведя. Оригинальная раскраска животного умиляет и...
В труднодоступных лесах на западном побережье Канады живет таинственный медведь-призрак, как назвали его местные индейцы. Его впервые описал Френсис Кермод, поэтому медведь называется...
Панды – милые и красивые черно-белые мишки, ареалом обитания является территория Китая. Название панды с китайского переводится как, «медведь кошка». Публика Западной Европу...
Конечно, вопрос о существовании медведей в огромном мегаполисе можно считать глупым, но есть же ещё и область. И, по последним данным, многие медведи...
Рацион уссурийского белогрудого, или гималайского, медведя достаточно разнообразен и во многом определяется ареалом его обитания. Так, медведи, обитающие в тропических лесах, являются хищниками,...
Панда относится к хищным животным, а основная пища составляет растительность, и по большому счету их можно назвать вегетарианцами. 99% их еды составляет бамбук....
Практически каждый рыбак мечтает о том, чтобы выловить большую рыбину всем своим коллегам на удивление. Когда заходит разговор о том, какие экземпляры попадались...
Дрейфующие и припайные ледники являются местом обитания белых медведей. Именно на льдинах они производят охоту и добывают основную часть своего пропитания. Рацион белого...
Каждый человек в своей жизни желает увидеть разнообразных животных в естественной среде обитания. Но обычным городским жителям очень тяжело осуществить такую мечту. Для...
Кадьяк – это подвид бурого медведь, данное животное относится к семейству медвежьих. Этот зверь имеет огромные размеры и зачастую его можно сравнить с...
Черный медведь или барибал (Ursusamericanus) принадлежит к млекопитающим семейства медведей – самых крупных представителей отряда млекопитающих хищников. Этот вид медведей проживает на землях...
Черный медведь имеет еще одно название – барибал. Это животное имеет множество внешних отличий от бурого медведя и образует отдельный вид. Проживает барибал...
Очень забавное и смешное животное – панда. Живет такое чудо в листвяных и хвойных лесах с хорошей бамбуковой прослойкой в центральных областях Китая....
Названий у гималайского медведя несколько. Иногда можно услышать белогрудый медведь, черный азиатский медведь или же лунный медведь, но все это относится к одному...
Зачастую панды – это молчаливые животные, они любят тишину. Но иногда все-таки можно услышать некий звук, похожий на блеяние козлят или ягнят. Такой...
Медведей невозможно перепутать ни с кем, хотя медвежьи виды разнообразны. Они различаются по внешним и анатомическим характеристикам, местам распространения и образу жизни. Их...
Медведь – это хищное крупное млекопитающее, относится к подотряду псообразных. Отличительными чертами являются коренастое сложение тела, мощные когти и клыки, короткий хвост, густая...
Медведи являются одним из интереснейших видов на планете. Практически у каждого человека, увидевшего медвежонка, возникает желание погладить и поиграть с ним. При общении...
Еще 300 лет назад панды населяли почти всю территорию Китая. Большинство их популяции находилось на заснеженных территориях Тибетских гор. Черно-белый окрас помогал медведям...
Медведь гризли входит в десятку самых опасных существ на планете. По размерам он значительно превосходит своих европейских братьев. При весе 500 кг, его...
Каких только в природе нет животных. У одного вида только такое большое разнообразие по структуре, внешнему виду и поведению. К примеру, медведи бывают...
Панды являются млекопитающими и относятся к семейству енотовых, хотя внешне очень похожи на медведей. Однако отличается от обычных медведей более длинным хвостом, формой...
Основной отличительной чертой азиатского черного медведя является V образное белое пятно в области груди. Именно поэтому в некоторых регионах его называют «Лунный медведь»....
Этот крупный хищник обитает в северном полушарии в границах дрейфующих льдов Арктики, этот близкий родственник бурого медведя настолько преобразился в суровых условиях севера,...
Типичным представителем фауны арктических широт является белый северный медведь – крупнейший хищник из своего семейства. Почти тонну веса имеет взрослый самец, а длина...
Эти милые животные радуют всех своим лучезарным и прекрасным видом, они похожи на мягкую игрушку, которую хочется потискать и обнять. Их привычная среда...
Когда берешь в руки декоративного кролика, получаешь ощущение, что в руках мягкая игрушка. Маленькие — они очень потешные, подростая, становятся еще и шалунами....
В нашем государстве при слове «медведь» у всех возникает ассоциация с бурым медведем. Но в природе, кроме бурого медведя, существуют еще такие виды:...
Одним из самых красивейших животных, семейства медвежьих, является гималайский медведь. Очень часто медведя можно встретить на разных картинах. Его отличает белый треугольник на...
Белые (полярные) медведи в отличие от своих собратьев, обитающих на суше, питаются исключительно тем, что добудут на охоте: это хищники, причём самые крупные...
Медведи являются дикими животными, но, тем не менее, многие пытаются приручить их, посадив в вольеры. Для дикого животного помещение в неестественную для него...
Медведь гризли представляет собой подвид бурого медведя, однако, в отличие от него, обитает он на территории Северной Америки и Канады в изолированных зонах...
Белых медведей называют полярными за счет того, что они обитают на приполярных морских льдинах Арктики. Ледяные просторы являются их родным домом. Белый медведь...
Черных медведей, или барибалов, часто называют симпатичными. Они имеют репутацию глупых животных, однако это совсем не так. Эти медведи выступают настоящими хозяевами своей...
Барибал – обитатель северной части Америки. Является территориальным соперником бурого медведя, часто называемого североамериканским. Однако значительно уступает ему в размерах – всего 1,5...
Опытные дрессировщики считают медведя одним из самых опасных хищников. Его поведение, в отличие от других животных, предугадать невозможно. Изменение настроения представителей семейства кошачьих,...
Белый медведь в наше время является самым большим представителем хищников на Земле. Живет на берегу Северного Ледовитого океана и во льдах Арктики, насчитывается...
Медведь — могучий и уважаемый сосед человека с самых давних времен. На Дальнем Востоке проживают дикие, практически отчужденные от человека камчатские медведи. Эти...
Отличительной особенностью белого медведя является прожорливость. Белые медведи на сегодняшний день являются самыми крупными представителями животного мира. Взрослые самцы не редко достигают до...
Барибал, или американский чёрный медведь— млекопитающее из семейства Медведи. Некоторые выделяют его в род Euarctos. В действительности это обычный медведь, который распространён от...
Не все представители человечества осознают разрушительный эффект глобального потепления. Некоторые предпочитают вовсе не замечать его, называя системные климатические изменения обыкновенным потеплением или похолоданием....
Оригинальное животное, которое можно узнать сразу, это медведь. У большинства представителей этого вида, туловище небольшое, толстое, медвежата могут быть стройными. Голова у медведей...
Эти сильные и крупные животные всегда привлекали к себе много внимания. Разные культуры восхищались медведями, поклонялись им и пытались перенять некоторые их повадки,...
Ареал обитания полярных медведей ограничен зоной Арктики. Здесь самые большие представители семейства хищных имеют все самое необходимое для выживания: условия постоянных холодов и...
Семейство медвежьих представляют самые крупные хищники, живущие на нашей планете. Эти огромные млекопитающие умеют не только стоять на задних лапах, но и проходить...
Медведь – это удивительное создание. Он умеет лазить по деревьям, хорошо плавает, быстро бегает. Эти животные всегда вызывают восторг у людей. А где...
Люди называют медведя косолапым не совсем справедливо. Лапы у животного совсем не кривые. Скорее его можно назвать «Иноходцем». Так шагают породистее лошади. Попеременно...
Многие люди ходят в зоопарки, чтобы посмотреть на самого большого сухопутного хищника, который привлекает своими повадками как детей, так и взрослых. А некоторые...
«Столбы» — это туристско-экскурсионный заповедник, который предлагает своим посетителям обширную программу с посещением наиболее красивых и примечательных мест. Походы в заповеднике стали излюбленным...
Преддипломная или производственная практика – это время подготовки практической части дипломного проекта, важная часть учебного процесса, завершающий этап обучения в ВУЗе, во время...
Жизнь полна неожиданностей и сюрпризов. К сожалению, некоторые из них носят и не совсем приятный характер, что отображается на нашем эмоциональном состоянии и...
Проблема электризации волос волнует многих, ведь от хорошей прически меняется не только внешность, но и настроение. А если волосы сильно электризуются, то, после...
Выбирая детскую обувь, нужно ориентироваться на ее удобство, экологичность и легкость. Мокасины прекрасно сочетают в себе все перечисленные достоинства. Независимо от возраста ребенка,...
В настоящее время выпускается огромный ассортимент всевозможной мебели для гостиной. Ведь гостиная — это не простая большая комната в квартире, а то место,...
Главными прелестями курортного маленького городка Анапа являются жаркий климат, целебные грязи и минеральные родники. Провести отдых в этих удивительных краях будет замечательным подарком...
«Золотой бубен» — фестиваль телевизионных фильмов и программ, созданный в 1999 году по инициативе губернатора Ханты-Мансийского автономного округа – Югры. В этом открытом...
Можно назвать это заболевание распространенным. Эпителиальный копчиковый ход имеет множество иных названий: киста или свищ копчика и др. Дело в том, что под...
После рождения малыша, многие молодые мамы отказываются от мини юбок, и сменяют их на более удобную повседневную одежду, отказываясь от каблуков в пользу...
Сувенир, как памятная вещь, приобретённая в подарок, должен быть оригинальным, напоминать о событии, памятной дате, путешествии, проведённом вдвоём или с друзьями. Зачастую сувениры...
Для многих крупных производителей, изготовление кормов для животных стало хорошим бизнесом, дающим многомиллионную прибыль. В сложившихся рамках уже никого не интересует здоровье домашних...
Панды сегодня – чуть ли не самые популярные животные. Панды – это практически настоящие звезды животного мира! А потому вдвойне печальнее осознавать, что эти...
Ни для кого не секрет, что для максимальной устойчивости организма человека к неблагоприятным условиям внешней среды нужно его укреплять. Самым распространенным и общеизвестным...
Подбирая тренажер, потребитель зачастую хочет приобрести устройство, которое помогло бы и здоровье укрепить, и сбросить лишние килограммы. Но при этом, чтобы не были...
Для любительниц классики и гламура существует куча всяких сумочек: больших и маленьких, ярких и нейтральных тонов, с разными узорами и украшениями. А вот...
О медведях существует множество сказок и легенд. В любой местности, где жили и живут медведи, обязательно рассказывают какие-нибудь истории. Есть люди, которые всерьёз...
Приближается наш давно ожидаемый отпуск, а мы до сих пор не знаем, какую страну или курорт выбрать для отдыха. Веб-пространство пестрит яркими картинками...
Еще в рукописи Евклида «Начала», которая датирована ХVI веком, можно увидеть иллюстрацию, где изображена женщина, учащая своих детей основам геометрии. Хотя считается, что...
Во все времена, владение дополнительными навыками и знаниями всегда являлось несомненным плюсом. В наше время, благодаря развитию интернета, можно обучаться даже не выходя...
С давних пор человечество пыталось научить животных разговаривать на человеческом языке. Как правило, в 99% случаев ничего не получалось. Но ведь остается один...
Правильная работа любой фирмы или предприятия без выполнения требований по охране труда просто немыслима. От числа сотрудников, работающих в конкретной компании, и типов...
Если вы стремитесь начать свое дело и умеете работать сварщиком, знаете все тонкости этого дела, то вам следует развивать у себя мышление предпринимателя....
В современном обществе с понятием «прибыль» знакомы как школьники, так и пенсионеры. У предпринимателей же в прибыли, как таковой, заключен весь смысл их...
Создание интересных и гармоничных фотоснимков – настоящее искусство. Пожалуй, фотографирование животных освоить сложнее всего, ведь животные не позируют своим фотографам, настороженно воспринимают их...
Эймериоз крупного рогатого скота вызывается кокцидиями, паразитирующими в эпителиальных клетках слизистой оболочки толстого кишечника, а также 12-перстной и тощей кишок. Наиболее подвержен этому...
Природа России достаточно разнообразна, а как может быть иначе, если страна имеют большую протяженность с юга на север. За огромные площади леса Россию...
Бытовая техника является для современного человека является практически вещью первой необходимости, поэтому к ее выбору стоит подходить тщательно и не полагаясь на удачу....
Во всем мире огромную популярность начинает набирать испанский язык, как второй иностранный язык. Многим сотрудникам в большинстве крупных и известных компаний требуются знания...
Отдых в Турции – это незабываемое наслаждение, которое возможно получить от ласкового субтропического климата, хорошего пляжного отдыха, богатого культурного наследия и восточного колорита....
Эти крупнейшие наземные хищники сегодня обитают в Европе, на северо-западе Африки, в Азии, Арктике, Северной Америке и в горах Южной Америки. Самыми большими...
Для необходимой ориентации в теоретических, а так же практических подходах к методам обучения персонала, можно рассмотреть опыт некоторых компаний. Сделав соответственные выводы, вы...
Перманентный макияж — очень молодая и активно развивающаяся область косметологии, направленная на улучшение внешности. В любом возрасте женщина должна быть красивой, молодой и...
Для развития бизнеса приходится иметь дело с заграничными партнерами. Но для ведения с ними дел необходимо переводить множество документов с иностранного языка. К...
Студенчество – счастливая и в то же время напряженная пора, особенно во время приближения контрольных проверок знаний, экзаменационных сессий, защиты курсовых и, конечно...
Шкура медведя – дорогой и шикарный подарок, который придется по душе любому мужчине. Еще с древних времен она олицетворяла власть, богатство, материальный достаток,...
Белый медведь является самым крупным хищником из разряда млекопитающее, которое в свою очередь принадлежит к семейству медвежьих. При росте в полтора метра, масса...
Если у навахо, апачей, пуэбло и некоторых других народов, преимущественно на Юго-Западе страны и в тихоокеанских штатах, шаманство сохраняется в виде более или...
Столица России по праву считается обладательницей лучших университетов и институтов, а академическое образование ценится и за пределами Российской Федерации. Чтобы гарантированно получить высокооплачиваемую...
Реалии современного времени таковы, что многим людям для успешной карьеры необходимо владеть английским языком. В этих случаях, проблема его изучения в максимально сжатые...
Анапа – курортный город, который ежегодно встречает большое количество туристов во всех стран мира. Анапа считается райским уголком для семейного отдыха. Для туристов...
Трансфером считается специальная транспортная услуга, с помощью которой можно доставить пассажира и его багаж на заранее заказанном автомобиле с водителем. Заказывать авто можно...
Курсовая – особая работа, назначение которой – проверка степени усвоения студентом материала по какому-то из изучаемых предметов в течение учебного года. Выполняется и...
Когда у ваших питомцев, а в частности собак, неожиданно возникает истощение, связанное с нехваткой минеральных компонентов, а также нарушается обмен веществ, то вашему...
Первое знакомство с бизнес-образованием или MBA произошло ещё в начале 90-х годов.Но тогда к этому относились скептически.Бизнесмены не понимали зачем им это нужно.Недоверие...
На последних курсах университета пред студентами встаёт вопрос о написании дипломной работы. Это не такая лёгкая задача, как кажется на первый взгляд. Без...
Анапа – курорт, на который каждый год приезжают миллионы туристов. Поэтому на его территории находится множество санаториев, пансионатов, развлекательных учреждений. Большинство людей работают...
Для претворения идеи о постройке дома в жизнь, необходимо приобретение участка и получение в местной архитектуре разрешения на постройку дома. Для получения разрешения...
Покупка товаров для школы – дело непростое и хлопотное. Конечно, родители первоклассников идут в магазин, вооружившись внушительным списком. Однако мамы и папы подросших...
Курортный город Анапа не просто получила звание популярного детского, а также семейного отдыха. Ведь тут можно не только греться в лучах солнышка, купаться...
Отношение людей к охоте очень неоднозначное. Для кого-то это настоящее варварство, другие же находят в этом удовольствие. Почему так происходит? Попробуем разобраться. Что...
Город Арзамас является одним из самых удивительных мест. Он входит в исторические города России. Это именно то место где, со старыми памятниками архитектуры...
В мире существует общепринятая классификация гостиниц по пятизвездочной шкале. Гостиницы, предоставляющие наиболее низкое качество услуг, оцениваются в одну звезду, наилучшие гостиницы получают пять...
Нередко, недовольные своей внешностью граждане, начинают задумываться о своем чудесном перевоплощении после пластической хирургии. Но, если некоторые обижены на природу, что та их...
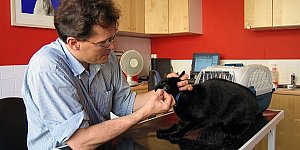
Ветеринария имеет очень долгую историю, зародилась она в деревне, благодаря вниманию крестьян к здоровью своего скота. Профессия «ветеринар» безусловно, является одной из самых...
Сокровищница мировой литературы содержит немало произведений классиков, которые были написаны века или даже тысячелетия назад. Что же такого ценного в этих произведениях, и...
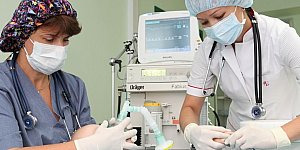
Животные, содержащиеся в зоопарках и специальных парках, требуют особенных условий для жизни, постоянной заботы о питании и здоровье. Поэтому люди, которые работают с...
В последнее время все большее число людей предпочитают проводить культурный отдых в зоопарке или отправляются на представление в цирк. Если в ближайшем прошлом...
В чем заключается психология медведя? Сегодня в зоопарке или цирке можно часто повстречать настоящего медведя, который имеет более добрый нрав, нежели дикие особи...
В отличие от множества других животных, медведи не создают постоянных пар. Как правило, пара существует вместе от четырех до двенадцати дней, после чего...
Недаром говорят, что ночная охота на медведя является самым опасным видом спорта. Дело в том, что именно ночью практически все виды медведей проявляют...
Медведи – крупные млекопитающие, обитающие по всей территории земли, начиная с крайних северных широт и заканчивая африканскими просторами. Весной или в начале лета...
Одной из неотъемлемых частей нашей жизни является дарение подарков своим близким, как с поводом, так и просто, чтобы сделать приятно. На каждый праздник...
Бурые медведи одни из самых известных с давних времён людям животных. Совсем не случайно изображение медведя встречается на гербах многих крупных городов, как...
Медведь губач – это млекопитающий хищник. Внешность мишки подвергла людей дать ему второе название – Ленивец. Но его облик обманчив. Этот зверь довольно...
К сожалению, во многих местах еще недавнего проживания в Европе, популяция этого известного под именем бурый медведь хищника резко сократилась. Человеческая деятельность сокращает...
В последнее время большую популярность приобретает экотуризм. Многие спешат на охоту или рыбалку. Очень часто, случается так, что лесу, в котором вы остановились,...
Медведь – один из самых знаменитых индивидуалистов в животном мире. Им чуждо такое явление, как стая; каждое животное держится особняком. Единственное исключение –...
Однажды труппа известного советского дрессировщика медведей Валентина Филатова гастролировала в Штутгарте (Западная Германия). Во время репетиции, которая проходила в спортивном зале, временно переделанном...
Тайваньский черный медведь с характерным белым галстуком на груди является эндемичным подвидом гималайского медведя на Тайвани — острове в Восточной Азии. В Азии...
Бурый медведь (Ursus ARCTOS) является наиболее распространенным видом семьи медвежьих, который обитает в Европе, Азии и Северной Америке.С научной точки зрения, больше всего...
Рацион медведей видоизменяется в зависимости от сезона и урожая различных растений. После зимней спячки пищей для медведя могут стать муравьи, осиновые побеги и...
Вообще медведь считается всеядным животным, но большую часть его рациона составляет растительная пища. Такую пищу, он поедает месяцами, ест практически все, всходы ржи,...
На юго-западе Германии начали активно приглашать туристов в новые большие парки развлечений. Так администрация по туризму земли Баден-Вюртемберг заявила, что на их территории...
Президент США Теодор Рузвельт в далёком 1902 году на охоте отказался стрелять в загнанного собаками, полузабитого и привязанного к дереву медведя. Он отказался...
Как уже было отмечено, механизм ориентации белого медведя практически не изучен, можно лишь отметить поразительную способность зверей определяться по странам света. Одним из...
Изображение медведей на любом материале всегда впечатляло и ухватывало взор проходящих мимо, ведь медведи — цари леса, с одной стороны милые и пушистые,...
Как уже отмечалось, белые медведи предпочитают держаться среди плавучих, но не мелкобитых льдов, перемежающихся участками открытой воды, т. е. у кромки льда, полыней...
Хотя она и не имеет определенных границ, но наиболее обширна. В подавляющем большинстве случаев звери попадают в районы, расположенные вне области их постоянного...
В основном приурочена к материковому склону Северного Ледовитого океана и на севере, по-видимому, совпадает с его границами, что обусловлено гидрологическими и гидробиологическими причинами....
Глаза у медведя небольшие, светло- или желто-карие за счет коричневой радужиньУ Величина бинокулярного зрения достаточно большая, около 180 °. В то же время...
Мочеполовая система белого медведя изучена недостаточно. Известно, что тестикулы сильно изменяются по величине в течение года, от октября — ноября (период покоя) до...
Год выпуска: 2004 Страна: США Жанр: Документальный Продолжительность: 53:14 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Стелла Ча, Эм Мэри Хаммерс Описание: На...
Год выпуска: 2005 Страна: Норвегия Жанр: Научно-документальный Продолжительность: 00:46:50 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Nat Geo Wild Описание: Северный Полярный круг.Сотни...
Год выпуска: 2005 Страна: Великобритания Жанр: Документальный Продолжительность: 00:47:13 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Saint-Thomas Production для Animal Planet Описание: Белые...
Посткраниальный скелет белого медведя также отличается рядом особенностей от скелета бурого медведя. По Верещагину, шейные позвонки относительно длиннее по оси, атлант с сильно...
Год выпуска: 1992 Страна: Великобритания Жанр: документальный Продолжительность: 6 фильмов по 49 мин. Перевод: Профессиональный (одноголосый) Русские субтитры: Нет Описание: Знаменитый сериал ВВС,...
Год выпуска: 2001 Страна: Чехия. Жанр: Про жив тных для детей. Продолжительность: 01:00:00 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Вацлав Чалоупек. Описание:...
По современным представлениям, родословное древо семейства медведей — Ursidae начинается со среднего миоцена от крупных представителей рода Ursavus, известного по находкам в Европе...
Бурый медведь (Ursus arctos). Внешний вид этого огромного зверя настолько хорошо известен, что нет нужды его описывать. Несмотря на усиленную охоту, до сих...
Год выпуска: 2004 Страна: Великобритания Жанр: Научно-популярный Продолжительность: 00:52:04 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Стив Банс / Steve Barnes Описание: C...
Подумать только, каким испытаниям подвергает иногда матушка-природа свои создания. Знакомясь с образом жизни некоторых зверей, невольно задаешься вопросом: «Как же они выживают?» Ведь...
Медведи, или Ursidae, — это тяжеловесы среди хищников. Аляскинский бурый медведь в расцвете сил или белый медведь, встав на задние лапы, достигает в...
Малайского медведя можно смело называть самым древесным видом из всех медведей. Благодаря своим длинным серповидным когтям на всех четырех лапах они прекрасно лазят...
Глобальное потепление, это очень опасно!!! Особенно для животных, которые привыкли жить в вечной мерзлоте. На сегодняшний день учёные и экологи все чаще бьют...
Наверняка Вы часто задавали себе вопрос, а почему именно так? Сейчас Вы узнаете, самые интересные факты из жизни медведей! Если хотите быстро определить...
Этот мишка является моногамным, – он не склонен к беспорядочным связям, а строит «серьезные отношения». Брачного сезона у медвежьих пигмеев не наблюдается: свадьбу...
Уссурийский медведь – типичный медведь. Как и хорошо нам известный бурый, он плотного сложения, неуклюж. На толстых и сильных ногах. Когда ходит, немного...
Бу?рый медве?дь (лат. Ursus arctos) — хищное млекопитающее семейства медвежьих; один из самых крупных наземных хищников. Бурый косолапый мишка — лесное животное. Обычные...
Очковый медведь (Tremarctos ornatus) , второе название коротколицый, единственный из семейства встречающийся в фауне Южной Америки. Населяет горные регионы (до 3000 м, был...
Белый медведь, является самым крупным наземным представителем млекопитающих отряда хищных. Его длина около 3 м, масса может быть до 1000 кг. Главное отличие...
Малайский косолапый мишка (Helarctos malayanus), или бируанг, обитает в тропических и субтропических лесах предгорий и гор Юго-Восточной Азии. Он распространен от северо-востока Индии...
Животные часто используются для изображений на логотипах и гербах. Не миновал этой участи и медведь. Спортклубы, политические партии, учебные заведения и другие компании...
Медвежий праздник начинают праздновать ночью. Ханты и манси верят, что именно ночью боги посещают землю. А значит, они могут увидеть, как почитается тень...
Определённо все медведи между собой схожи, различие состоит лишь в приспособленности к конкретной среде и внешнему виду. Белый тип не живёт в густонаселённых...
В тайге часто можно встретить хищника особо крупных размеров – медведя бурой окраски. У этих животных передние лапы обладают большой мощью. Своими когтями...
Гималайские медведи – обитатели субтропиков. Лесных массивов с высокими деревьями. Родина гималайского медведя тайга южной части Дальне востока в России. Гималайского медведя можно...
Белые медведи живут там, где белым-бело, на севере. По количеству северных пространств наше Отечество впереди планеты всей. Одна треть России лежит выше 60°...
Малайских медведей называют бируанги. Они являются самыми маленькими представителями среди своих родственников. Максимальный рост — полтора метра, а вес — 65 килограмм. В...
Медведи — обитатели исключительно Северного полушария. Среди млекопитающих они отнесены к отряду хищных. Это справедливо только отчасти, ведь большинство представителей этого семейства являются...
Этот представитель семейства медвежьих отличается острой мордой и небольшими размерами по сравнению с другими видами. Он обитает в Северной Америке. Его естественный ареал...
Все мы знаем этого замечательного мишку, который обитает в Азии. В последние века его причудливый черно-белый вид и добрые повадки вдохновляют многих авторов...
Медведи – они такие разные… Есть бурые, белые, черные, гималайские, белогрудые, гризли, а также многим полюбившиеся милые мишки панды. И на некоторых из...
Медведи – крупные млекопитающие, обитающие от Южной Америки до Арктики. Могут жить в различных климатических условиях от степей до вечных льдов. Насчитывается восемь...
Самыми большими хищниками на планете считаются белые медведи, которые обитают в Канаде, Норвегии, Гренландии, России и на Аляске. Большая часть животных находится в...
Герб столицы Германии обычно узнаваем по изображению на нем медведя. Некоторые исторические свидетельства подтверждают, что город когда-то был основан вблизи берлоги этого лесного...
Панда, или бамбуковый медведь – милое и симпатичное животное, отличающееся от остальных собратьев своего рода необычным окрасом. Хотя далеко не все зоологи относят...
Одними из самых умных и сообразительных зверей являются медведи. Они отлично дрессируются, быстро учатся новым командам. Несмотря на эти достоинства, для дрессировщика медведи...
Большая панда является неофициальным символом Китая. Данный вид медведя можно увидеть только здесь, так как нигде в мире больше нет таких уникальных природных...
Символ русского человека – бурый медведь. Большой, с виду неповоротливый, косолапый. А на деле – очень опасный хищник. Тот, кто недооценивает его неуклюжий...
Белый медведь прекрасно обходится без зимней спячки, в отличие от бурого собрата. И только беременные самки позволяют себе понежиться в снегу, в ожидании...
Мексиканский медведь гризли является подвидом бурого медведя и был официально признан вымершим в шестидесятых годах прошлого столетия. Это было довольно большое животное, самое...
Медведь входит в число наиболее опасных диких зверей. Если вы встретите его в природных условиях, то сможете получить очень необычный опыт. При этом...
Что же подарить охотнику? Незнающие люди перечислят множество ненужных подарков для охотника, которые в конечном итоге будут просто забыты, но знающие остановят свой...
Белый медведь относится к семейству медвежьих. Данное семейство появилось на свет более 5 миллионов лет назад. Обитают белые медведи в Америке и Евразии,...
Белый медведь имеет хорошо развитое зрение, обоняние и слух. Благодаря этому такие животные чуют добычу на достаточно большом расстоянии. Зачастую медведи ходят не...
Разновидностей медведей великое множество, географически они разбросаны по всей планете, от южного до северного полюса. Рассмотрим самые известные из них. Бурый или обыкновенный...
Белогрудого медведя еще называют черным уссурийским и черным гималайским медведем. Как правило, обитает такой медведь в горных и холмовых лесах, которые протянулись от...
Эндемиком островов Кадьякского архипелага является особый вид бурых медведей – кадьяк. Остров Кадьяк находится на юге Аляски, поэтому данных медведей часто называют аляскинскими...
Серый медведь имеет второе название «гризли». Такие медведи представляют собой подвид обычного бурого медведя, который признан самым крупным животным. Без компьютера сейчас немыслима...
Большая панда очень красивое, необычное и благородное животное, имеющее степенный нрав и обладающее рассудительностью. Недаром Китай выбрал ее символом своей страны и почитает...
Барибалов часто можно встретить в Северной Америке. Ареал их обитания невероятно велик: черные медведи встречаются в лесах от севера Аляски и Канады, до...
Широкой известностью и любовью среди всего населения земного шара пользуется такое животное как панда. Пожалуй, сегодня трудно встретить человека, который при виде милой...
Медовый медведь – это другое название малайского медведя. Из-за маленьких размеров местные жители прозвали его медведь-собака, при росте 150 см, их вес не...
Бурый медведь это самый большой наземный хищник в России, наиболее крупными считаются камчатские хищники, которые весят до 600 килограмм, и к слову, они...
Не все знают, что есть просто память и память ассоциативная, которая является основой творческой деятельности человека. И в повседневности люди никогда не задумываются...
Мир животных всегда привлекал и очаровывал своей красотой и оригинальностью, богатством и уникальным строением. Однако в наше время становится все больше и больше...
С детских лет мы все любим пушистых и милых медведей. Одним из самых красивых видов медведей, бесспорно, является большая панда. Природным местом обитания...
Панды относятся к семейству медвежьих, они получили название «бамбуковый медведь». Это звери с черно-белой шерсткой. Они комфортно себя чувствуют в природных условиях горного...
Вряд ли найдется еще одно животное, мнение о котором столь ошибочно, как о медведях. В русской культуре это глуповатый, добродушный увалень, вечно остающийся...
Бируанг – это малайский медведь – самый миниатюрный и самый агрессивный из представителей семейства медведей. Он имеет несколько имен: из-за его маленьких размеров...
Гризли считают подвидом обычного бурого медведя, но норова и силы в них гораздо больше, они выделяются своей свирепостью и жестокостью, желательно не попадаться...
Среди многочисленных видов медведей умиление вызывают панды. Эти черно-белые неуклюжие животные с «очками» на мордочке пользуются популярностью в зоопарках. Однако, мало кому известно,...
Медведи, эти большие и мохнатые создания, кажутся нам столь неуклюжими, что даже мысль об их умении плавать представляется нам невероятной. Тем не менее,...
Как правило, к среде для обитания большой панды, принято относить: гористые участки Тибет-Цинхайского плато, западную часть гор на территории. Большая панда приспособилась к...
Природные ресурсы, за счет которых живет человечество, постепенно истощаются. Сегодня настал такой момент, что все развитые страны обращают свое внимание на Арктику. Именно...
Бурый медведь – хозяин леса. Он обитает в густых непроходимых лесах горных местностей. Жизнь бурого медведя самца протекает в одиночестве. Он подходит к...
Как показывают опросы, многие люди даже не представляют, как выглядят и кто такие малайские медведи. Данная несправедливость будет исправлена и каждый, кто прочтёт...
Большую панду часто называют бамбуковым медведем, так как питается она исключительно бамбуком. От остальных медведей большая панда отличается пятнистым окрасом, а так же...
Малайский медведь является самым миниатюрным представителем медвежьих. Второе его название- бируанг. Основное место обитания этого вида располагается в субтропических лесах от Индонезии до...
Большие панды — симпатичные и добродушные животные семейства енотовых, обитающие в непроходимых зарослях бамбука в Индокитае и Китае. Они употребляют в пищу молодые...
Панда, или, как еще принято называть такой вид – бамбуковый медведь, является одним из особей семейства медвежьих, которые отличаются своей своеобразной черно-белой окраской....
Лечение домашних животных процедура простая и вполне обычная. В специализированных клиниках врачи быстро окажут необходимую помощь кошке или собаке. Лечение диких животных и...
На первый взгляд бурый медведь может показаться опасным хищником, но на самом деле ежедневный рацион лесного зверя состоит исключительно из растительных ингредиентов. Бурые...
Медведи – это крупные, хищные звери, которые представляют большую опасность для жизни и здоровья человека. Правила поведения в среде обитания медведей Любому любителю...
Очковый медведь – это южноамериканский медведь, занесенный в Красную книгу, так как находится на грани вымирания. Этот зверь небольшой величины, с белыми или...
Столь дружелюбные персонажи из детских сказок медведи запоминаются как добрые, миролюбивые звери. И правда, с лисьей хитростью и волчьей кровожадностью они и рядом...
Чёрных медведей с белой летящей птицей на груди называют гималайскими медведями по месту их обитания. по-латыни название этой породы звучит несколько иначе: Selenarctos....
Медведи по праву называются самыми крупными и грозными хищниками на планете Земля. По своим размерам они превосходят даже львов и тигров. Именно благодаря...
Большая панда обычно ведет спокойный, уединенный образ жизни, прячась в густых бамбуковых лесах. Лишь весной, когда все вокруг начинает цвести, природный инстинкт заставляет...
Бурые медведи занимают большую территорию обитания – пустыни и леса, высокие горы и заснеженные области. Местом широкого их обитания когда-то были леса Европы,...
Совсем недавно многие леса Европы были местом обитания большого количества бурых медведей. Представители вида встречались во всех европейских регионах и достигали в своем...
Из века в век вокруг медведей образовался ореол некой таинственности. Ведь эти опасные хищники повадками походят на человека, но и для последнего остаются...
Во многих фольклорных произведениях разных народов мира медведи выступают в виде самого величественного персонажа. Ведь это животное во многих своих привычках и повадках...
Медведи относятся к отряду хищных млекопитающихся, и отличаются от других животных своим могучим видом и коренастым телосложением. Медведи передвигаются не только на четырех...
Знакомство детей медведями можно с того момента, когда ребенок способен фикусировать внимание. Мы рекомендуем начинать знакомство с годовалого возраста, но если вы хотите раньше...
При наступлении теплого времени года все больше и больше людей предпочитают отправляться в лес для отдыха. Но здесь обитает множество зверей, которые считают...
Если вы помните, то на на утренниках и в детских спектаклях, очень часто можно встретить актёра, изображающего медведя. Однако, сыграть медведя — настоящий...
Косметика для собак включает в себя огромное разнообразие всевозможных шампуней, спреев, муссов, гелей, антистатиков, пудры и прочих средств, которые делают наших четвероногих друзей...
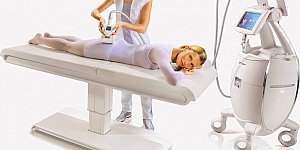
Массаж представляет собой различные приемы, с помощью которых происходит воздействие на органы и ткани человека с помощью давления, растирания или вибрации. Данные действия...
В наше время стало популярным изучение иностранных языков. Форм обучения языка существует большое количество, и выбрать правильное направление совсем непросто. Прежде чем приступить...
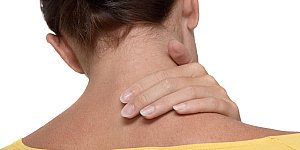
Стоит отметить, что остеохондроз шейного отдела по своим симптомам значительно отличается от симптомов, которые проявляются в других отделах позвоночника. В шейном отделе позвонки...
Одним из многих способов заработать деньги, не имея специальных умений и навыков, является работа курьером. На рынке труда вариантов этой вакансии великое множество....
Сегодня практически у каждого совершеннолетнего человека имеются кредиты. Банки охотно предоставляют займы своим клиентам и в результате у потребителей появляется возможность приобрести практически...
С развитием прогресса появляется масса новинок с огромными функциональными возможностями за умеренными ценами. Естественно, что мы с готовностью покупает разрекламированные товары, а вот...
Практически каждый взрослый человек окончил какое-либо высшее образовательное учреждение. Есть ли в нем смысл? Давайте разберемся. Современные университеты и Академии России готовы предложить...
Отношения с мужчиной порой теряют былую пылкость, накапливают взаимные обиды, легкость и комфорт сменяются непониманием. Конечно, восстановить отношения намного проще не тогда, когда...
Непознанный мир полярных медведей манит к себе не только фотографов-экстремалов, желающих заполучить эксклюзивные снимки, но и любителей адреналина. Ежегодно уничтожаются тысячи огромных хищников....
Черногория расположена в уникальном для отдыха уголке Европы, где из условий умеренно-континентального климата севера страны за несколько часов можно приехать в средиземноморский на...
Особенности природы Вследствие образования водохранилищ на Днепре значительно сократились площади, занятые луговой растительностью. После сооружения Кременчугской и Каневской ГЭС глубоким изменениям подверглись также...
В потоке повседневности приходится решать насущные проблемы различного характера, в списке которых часто отсутствует забота о здоровье и посещение врача. Неоспоримый факт —...
Освоение пространства и постижение окружающего мира требует от маленьких детей постоянной активной деятельности. Даже без руководства со стороны взрослого, маленькие дети многому учатся...
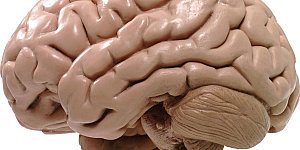
Ишемия мозга – недуг, вызванный локальной неспособностью участков мозга получать кислород. Обусловлено это недостаточным кровоснабжением коры головного мозга. Кислородное голодание происходит по причине...
Время бежит и дети подрастают, наступает момент, когда им нужно определить, куда пойти учиться. К этому вопросу необходимо подойти с ответственностью, так как...
Фотоискусство – дело не из простых. Может показаться, что приобретения хорошей профессиональной фотоаппаратуры достаточно для получения красивых и, главное, незабываемо впечатляющих фотографий. Это, безусловно,...
Муфлон — жвачное парнокопытное животное, относящееся к роду баранов. Эти животные могут достигать роста 90 сантиметров и веса 50 килограмм. В настоящее время...
В последние годы сократилось число россиян, которые проводят свой отпуск в Египте, Турции или Болгарии. Наши соотечественники стараются находить более экзотические страны, путешествуя...
То, что без знаний иностранного языка в сегодняшнем деловом мире делать нечего, уже все давно осознали. Посреднические услуги переводчиков не всегда являются оптимальным...
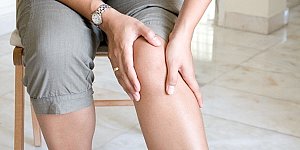
Коленный мениск чаще всего воспаляется при таких травмах, как чрезмерное разгибание и сгибание коленного сустава, ушиб голени тяжелым предметом, вывих колена, разрыв и...
Полимерные мешки завоевали огромную популярность во все мире, за достаточно короткий промежуток времени. В первую очередь подобное явление объясняется низкой себестоимость синтетического материала,...
Реферат – это повествование на какую-либо тему с использованием нескольких источников информации, так или иначе касающейся ее. Чаще всего такой вид работы дается...
В настоящее время, обучение за рубежом, стало очень популярным и престижным. Все больше и больше наших соотечественников, в особенности молодежь, стремятся выехать на...
В наше время не все желающие могут поступить в высшие учебные заведения и получить соответствующее образование на бесплатной основе. Бюджетных мест по каждой...
Доктор Айболит один из любимейших сказочных героев многих поколений. Практически все дети любят щенков и котят, птичек и рыбок. И, конечно же, многие,...
Еще не закончился учебный год, а родителей уже волнует вопрос «Как рационально спланировать отдых ребенка в каникулярное летнее время». Ведь за 3 месяца...
Знание любого иностранного языка, несомненно, пригодится человеку в жизни. Какие возможности открывает это знание! Общение с людьми из чужой, далекой страны, погружение в...
Каждый человек рано или поздно становится перед выбором, куда отправится отдыхать. Вариантов для современного человека очень много, но важно выбрать такой, который отвечал...
Выполняя ремонт, не стоит загромождать пространство многоуровневыми потолками с различными видами подсветки, а стоит проявить фантазию и перенести источники света на другие поверхности....
Ребенок дошкольного возраста уже имеет четкое представление, что означает слово «медведь». Он может безошибочно выбрать из множества картинок изображение медведя. К 5 годам...
На богатой лесами территории России, среди множества представителей медвежьих (иначе – медведевые) встречается несколько: Белый и Бурый медведи. Бурый медведь Представителей этого вида...
Согласно словарю Даля, эрудированный человек – ученый, начитанный человек, разносторонне образованный и обладающий знаниями в определенной научной отрасли. Жизненный опыт говорит нам, именно...
Каждый год многие из нас, сталкиваясь с проблемой выбора верхней зимней одежды, останавливаются на пуховике. Не удивительно, ведь правильно подобранный пуховик легок, комфортен...
Болгария – одна из стран, где русские туристы чувствуют себя легко и свободно, ведь языкового барьера там не существует, болгарский язык очень напоминает...
Основанием множества неудач в спорте считается нецелесообразное распределение в меню, невосполнимость энергетических расходов и неумелое применение стимулирующих средств. Поэтому правильный рацион — залог...
Известно, что медведи наносят на деревья метки – царапают кору когтями или зубами, трутся спиной о стволы, оставляя на них клочья шерсти. Существует...
Все чаще можно встретить человека, который задается мыслью, что пора бы освоить иностранный язык. Причин может быть много, кому-то необходимо это для работы,...
Итогом деятельности студента за год, а соответственно и оценка его компетентности, является курсовая работа, грамотно написанная и не менее грамотно защищенная перед коллегией....
Творчество – наиболее приятный и эффективный инструмент не только взаимодействия родителей с малышом, но и развития основных психических и физических процессов ребенка. Сборность...
Натуральные меха издавна ценились и почитались. Наличие меха в доме, указывало на достаток и уют в жилище, на удачливого и опытного хозяина, настоящего...
Джубга – небольшой курортный поселок городского типа, расположившийся на побережье Черного моря в Туапсинском районе (Краснодарский край). Большую популярность этот населенный пункт имеет...
Любое хирургическое вмешательство, как правило, сопровождается болевыми ощущениями для больного. Порой они настолько сильны, что без анестезии (обезболивания) операция невозможна. Поэтому еще с...
После распада Советского Союза возможности граждан касательно мест, где можно провести отпуск резко возросли. Теперь не проблема съездить в Египет к вековым пирамидам,...
Многие родители сталкиваются с просьбами ребенка завести домашнее животное. У детей может возникнуть желание завести домашнего питомца в любом возрасте и по разным...
Сегодня довольно часто у детей встречаются различные дефекты речи, и помочь их исправить может логопед. Все дети индивидуальны, но следует помнить, что детская...
Отдыхать на белоснежных пляжах, которые утопают в тропической зелени, всегда доставляет огромное удовольствие. Курорты Таиланда привлекают туристов дайвингом. Огромное количество центров по дайвингу...
Если вам хочется получить новогодний подарок лично от Санта-Клауса, то приезжайте в Финляндию. Здесь вы сможете познать всю радость зимнего отдыха: снегоходы и...
Эта великолепная страна всегда могла привлечь туристов с любого уголка нашей планеты, и она считается самым посещаемым государством на всей Европейской территории. Франция...
Из года в год число автомобилей на российских дорогах (а, значит, и водителей) растет. Окончив автошколу и получив долгожданные права, приобретя собственный автомобиль,...
Соль и специи (пряности и травы) ежедневная составляющая различных блюд нашего рациона. Однако прежде чем включить в рацион ребенка что-либо из данных ингредиентов,...
Как известно, нефть является одним из наиважнейших и самых значимых энергоресурсов для современного мира. Черное золото, а именно так называют нефтепродукты за их...
Выбрать рюкзак – дело совсем непростое, как кажется на первый взгляд. Как же правильно сделать выбор? Во-первых, он должен быть довольно вместительным, но...
В современном обществе множество различных товаров и услуг предоставляют иностранные агентства и компании. И все чаще и чаще приходится пользоваться переводом дополнительной информации...
Вы собрались поступать и уже сейчас задумались о том, как избежать переутомления? Оказывается, если развивать нужные навыки, закрепить уже имеющиеся знания и...
Дробовик относится к классу гладкоствольного огнестрельного оружия, позволяющего стрелять дробью или пулями. Как правило, стрельба из дробовика ведется с плеча, но возможны и...
Собираясь на рыбалку нужно учесть погодные условия, особенности водоема (какие виды рыб там обитают), и время пребывания. За основу возьмем рыбалку в теплое...
Любителям экологического туризма в поисках красивых природных мест для отдыха необязательно ехать далеко за Урал. Немало укромных и таинственных уголков сохранилось в Новгородской...
Правильное питание является одним из факторов, определяющих здоровье животных. Однако, кроме того, что корм надо купить, его следует и правильно хранить, чтобы корм...
Понятно, что любая хозяйка и хозяин хотят видеть свой дом уютным, удобным и стильным, потратив на его обустройство наиболее приемлемую сумму денежных средств....
Казалось бы, что написать курсовую или дипломную работу сегодня невероятно просто, ведь доступ в интернет есть практически у всех. Однако данное положение не...
Все медведи — животные достаточно выносливые. И какими-то специфическими болезнями практически не болеют. Однако, хоть и редко, но встречаются некоторые заболевания, которыми могут...
Неуклюжий, огромный, косолапый бурый медведь знаком большинству из нас по детским сказкам. Такое впечатление у большинства остается об этом обитателе таежных лесов на...
Медведь является символом Германии, именно это животное изображено на гербе Берлина. Bar в переводе с немецкого означает «медведь», отсюда название столицы государства. Фигурки...
Медведи – представляют семейство хищных псообразных млекопитающих с коротким хвостом, а также отличным слухом и обонянием. Эти животные всеядны, они прекрасно лазают, быстро...
Гималайский медведь по своим размерам немного меньше нашего бурого медведя. Есть отличия и во внешнем виде: голова у гималайского медведя имеет заострённую морду,...
Полярные медведи — это северные братья наших бурых медведей. Но они значительно крупнее всех остальных медведей: высотой они полтора, а длиной почти три...
Из всех представителей медвежьего семейства наиболее известным является медведь бурый обыкновенный. Длина его тела составляет более двух метров, а вес может переваливать за...
Медведи относятся к хищным животным. Они являются самыми крупными среди хищных зверей и достигают массы до семисот килограммов, а некоторые особи даже до...
А вы знаете, что канадские индейцы до сих пор считают бурых медведей человеческой расой. В доказательство этой теории они приводят множество фактов. Во-первых,...
Аляска — один из последних нетронутых уголков природы в мире. Зимы здесь долгие, на верхних склонах самой высокой северной горы Америки Маккинли зима...
Бурые медведи обладают, так называемыми бицепсами, мышцами, которые делают их сильными животными и которые позволяют им копать. Головы бурых медведей достаточно крупные, с...
Большинство самок начинают спариваться в возрасте четырех-пяти лет, а некоторые из них не задумываются о потомстве и до 25 лет. Многие медведицы оставляют...
Самый крупный наземный хищник – это, несомненно, бурый медведь. Вес самых крупных медведей достигает до 600 килограмм. Средний вес обычного бурого медведя около...
Пищевой рацион бурого медведя различен и больше зависит от года, какая урожайность кормовых культур. По выходу из берлоги медведь поедает муравьев, побеги осины,...
Агрессивными, как оказалось бывают не только люди, но и медведи. И более активна агрессия при встрече с человеком, когда медведь попадает в поселение,...
Что вы знаете о полярных медведях? Белые существа, живущие в холоде, не видящие ничего вокруг кроме обломков льда, снега, не чувствующие ничего кроме...
Что только не случается на белом свете, вот одна из таких историй. В данном качестве выступила медведица по кличке Девочка, которую дрессировал известный...
Еще до начала широких гидрологических исследований в Северном Ледовитом океане А. Педерсен высказывал предположение о том, что белые медведи вместе с дрейфующими льдами...
Принимая во внимание большую площадь ареала белого медведя, а также тот факт, что животные распределены здесь неравномерно и работы, связанные с их учетом,...
Границы областей постоянного современного обитания белых медведей и их высокой численности яются в зависимости от сезонов года, что связано с ледовым режимом Арктики....
В отличие от остальных особей вида, беременные медведицы имеют устойчивые связи с сушей, где осенью залегают в берлоги. Свои зимние убежища они чаще...
Область постоянного современного обитания животных. Не выходит за пределы зимней границы дрейфующих льдов в северном полушарии. Наиболее далеко (до 50 ° с. ш.)...
Для головного мозга белого медведя, в отличие от бурого, характерно более умеренное развитие передних долей больших полушарий, большее развитие зрительной области и наличие...
Уровень метаболизма у белого медведя, очевидно, выше, чем У бурого. У белого обнаружена также необычайная стойкость к воздействию низких температур не только за...
Температура тела взрослого белого медведя, измеренная ректально, составляет 36,8—38,8 °С (ниже, чем у бурого); суточные изменения температуры не отмечены. Температура поверхности кожи, измеренная...
Год выпуска: 2009 Страна: США — Великобритания Жанр: документальный Продолжительность: 0:49:01 Перевод: Профессиональный (полное дублирование) Русские субтитры: нет Режиссер: Роджер Джонс (продюсер) Описание:...
Линька у белого медведя происходит практически в течение всего года. Первыми с конца мая начинают линять взрослые самцы и годовалые медвежата; взрослые самки,...
Меховой покров взрослых зверей зимой очень густой, плотный, в отличие от меха бурого медведя более короткий, жесткий, с обилием остевых направляющих и промежуточных...
Печень белого медведя характеризуется необычайно высоким содержанием витамина А (до 9—10 тыс. мг/г и свыше 30 тыс. межд. ед/г), во много раз превосходящим...
Названия белого медведя на большинстве европейских языков происходят: от цвета меха зверя: бели медвед (сербско-хорв.), niedzwiedz bialy (польск.), l’ours blanc (фр.), orco bianco...
Сравнительно с остальными группами отряда хищных, представители семейства медвежьих отличаются наибольшим однообразием внешнего вида, размеров, многих особенностей внутреннего строения. Медведи — самые крупные...
Есть несколько островов в Ледовитом океане, которые белые медведи издавна облюбовали как места устройств «зимних квартир». В восточной части Арктики это архипелаг Земля...
Медведь медведю — рознь. Есть гиганты в два человеческих роста, тяжелые, живущие на открытых северных просторах. Среди них белый медведь. А есть и...
На малайского медведя охота больше ведется ради спортивного интереса и для продажи. Однако, известно что, в Китае части тела медведя стали использовать в...
Белому медведю, как никакому другому необходим жир, чтоб не замерзнуть. А для того, чтобы сохранять необходимый запас жира ему необходимо съедать очень много...
Размножение. Спаривающихся белых медведей можно увидеть весной или летом. Процесс очень интересный, около самки собирается от 3 и даже до 7 самцов. Брачной...
Может показаться странным, но кожа у белого медведя тёмного, можно сказать, почти чёрного цвета, это способствует наименьшей теплоотдаче. Почему тогда мех белого медведя...
Малая панда (Firefox) является родственником большой панды, и едва превосходит по размерам кошку. Своим рыжим окрасом и острой мордочкой, она напоминает лисицу. Было...
Гималайский косолапый медведь, он же белогрудый или уссурийский косолапый медведь (лат. Ursus thibetanus), млекопитающее отряда хищных. Гималайского медведя порой выделяют в отдельный род...
Медведь гризли (Ursus horribilis) Медведь гризли — это никто другой, как североамериканский подвид обыкновенного бурого медведя. На сегодняшний день он входит в десятку...
Белогрудого медведя (Ursus thibetanus), называемого также гималайским, порой выделяют в отдельный род Selenarctos, что означает «лунный медведь». Он обитает в холмовых и горных...
В Украине медведей можно встретить в лесах Карпат. Также их много в Закарпатье. В редких случаях одиночные медведи могут попасть из Белорусских лесов...
Его любят, уважают, боятся, слагают о нем сказки, песни, легенды. В сказках медведь обычно добрый и покладистый, но если верить древним преданиям, не...
Медведь — это крупное хищное животное класса млекопитающие. Он появился на планете примерно шесть миллионов лет назад и всегда являлся представителем мощи и...
Белыми медведями называют самых больших хищников, которые живот на поверхности земли. В природе можно встретить их 15 видов, а общее количество насчитывает примерно...
Гималайский медведь (белогрудый, черный) проживает на восточных и южных территориях, относится к отряду хищных млекопитающих. Данный вид довольно-таки распространен, но некоторые подвиды на...
Существует множество историй и слухов о редких животных нашей планеты и их способностях. Можно встретить и лигра, и щелезуба кубинского, и нарвала, и...
В мире существует немалое количество разновидностей медведей. Однако популярными являются лишь несколько видов, такие как бурые медведи, белые, медведи гризли. При этом очень...
Каждый род семейства медвежьих имеет свои особенности и по своему уникален. Среди них находится представитель, внешность которого не только примечательна, но и имеет...
Барибал – это медведь из мест северной Америки. Чаще всего встречается другое название – черный медведь. Привычные места обитания простираются от северной части...
Панда — является разновидностью медвежьего рода. Однако, она сильно отличается от своих предков. Учёные выяснили, что гены человеческие и панды одинаковы на 68%,...
Сотни лет назад видов медведей на Земле было значительно больше, чем нам известно сегодня. Одним из представителей семейства медвежачьих является гималайский медведь, так...
Многие удивятся: на планете есть город, в котором проживают 6 миллионов итальянцев. Не менее удивительно, что речь идет о крупнейшем мегаполисе Бразилии –...
Белых медведей еще называют полярный или северный медведь, а также есть и более научные названия: ошкуй, нанук, умка. Их среда обитания – дрейфующие...
Тем, кто знает эти фильмы, особенно щемящую душу работу французского режиссера «Медведь», не могли не запомнить огромного бурого зверя. Назван этот вид в...
Медведей издавна ассоциировали с магией и огромной энергетикой. Также в древности тех людей, которые не испытывали страха перед этими зверями, считали сверхлюдьми. А...
Медведь размером с крупную кошку – кажется, такое бывает только в мультфильмах. Однако такое живое чудо действительно существует в Юго-Восточной Азии, поражая зоологов...
На Западе Россия давно прочно ассоциируется с медведем. Хотя в мире существует большое количество видов семейства медвежьих, проживающих в самых разнообразных климатических условиях...
Неприметная деревушка Лакхапама, затерявшаяся на побережье Бенгальского залива в штате Орисса (Индия) оказалась ненадолго в центре забавной истории. Дело в том, что семейство...
В настоящее время медведи являются самым крупными представителями отряда хищников. Существует семь отдельных видов медведей. Эти виды отличаются друг от друга и среда...
Медведь это по истине величественный зверь. Является самым крупным наземным хищником. Во многих странах медведь представляет собой символ силы и мудрости. Самые распространённые...
У южных берегов Аляски на островах Кадьякского архипелага живет самый крупный сухопутный хищник — бурый медведь, названный по имени одного из островов, тоже...
Самый маленький из всех медведей — малайский, или бируанг — живет в тропических лесах и горах юго-западной Азии, на островах вокруг Южно-Китайского моря....
Белый медведь – хищник не робкого десятка. Кажется именно для него, созданы арктические просторы. Это невероятно хитрое и ловкое животное. Если, из –...
Являясь подвидом бурого медведя гризли, относится к более крупной и тяжелой породе. Шерсть его имеет легкий серебристый оттенок. Любитель вкусно и много поесть,...
С первых дней своего появления черный медведь проживал на территории Северной Америки. Зачастую данное животное выбирало равнины и леса. В конце IX века...
На данный момент пещерный медведь является вымершим видом семейства медвежьих. Его рождение было зафиксировано более 300 тысяч лет назад, а исчезновение – более...
Малайский медведь (Helarctos malayanus) – наименьший представитель семейства медвежьих. Он также известен как солнечный медведь, бируанг, медведь-собака, медовй медведь. Популяция этого вида встречается...
В культуре Китая гималайский медведь имеет особое значение как культурного, так и мифологического наследия. Его называют лунным медведем. Он олицетворяет мужество, силу, достоинство...
Большие панды — это млекопитающие, которые относятся к семейству медвежих. На родине панды — в Китае ее называют медведем-кошкой. Взрослая панда, в ширину...
Белогрудые медведи занимают довольно обширную территорию. Их почти столько же сколько простых бурых медведей. Данный вид в каждой месте обитания называют по-разному: черные,...
Полярные медведи самые крупные плотоядные существа на суше. Они населяют северные побережья Евразии и Канады. Среди вечных снегов Арктики обитают около 40000 белых...
Гималайский медведь – млекопитающее из отряда хищных, также имеет другое название – лунный медведь, из-за желтого пятна в виде полумесяца на его груди....
Во времена появления коал в Австралии подавляющую часть материка занимали эвкалиптовые леса, и природа создала живое существо, которое заняло пустующую нишу потребителя этой...
«Мети» – именно так называют гималайского медведя на родине в Тибете. Существует много мнений, что следы именно этого животного породили слухи о Йети...
Медведи являются плотоядными животными, ведущими довольно активный образ жизни. Поэтому для их содержания в неволе требуется выполнение специальных требований, ведь место их постоянного...
Медведи всего мира подразделяются на семь разновидностей. Названия виды в большинстве своем получили от окраса, только в случае совпадения оттенков, как это произошло...
Белые медведи являются одним из самых крупных видов хищных млекопитающих. От других разновидностей медведей его отличает длинная шея, плоская голова и, конечно, белая,...
Разнообразие животного мира огромно! Медведь, самый большой хищник на суше. Мощное животное, достигающее более полутора метров в длину. Вес этого грозного зверя может...
Основным местом обитания медведя считается Борнео, Ява, Суматра и Малаккский полуостров, где природная зона субтропиков и горных лесов считается самой подходящей для его...
Медведь является находчивым и сообразительным хищником, который обладает обостренным слухом и обонянием, но при этом, у него слабо развито зрение, особенно на дальние...
В медвежьем царстве, которое у нас, в первую очередь, ассоциируется с могучими и сильными бурыми медведями, их северными собратьями и медведями гризли, самым...
Самыми популярными животными в зоопарках всего мира считаются белые медведи. Взрослые и детки с удовольствием наблюдают за смешными и неуклюжими движениями этих замечательных...
Медведи являются интересными животными, несмотря на свой большой вес, легко способны взобраться на дерево, носиться галопом. Они относятся к хищным животным, но из-за...
В известной сказке Киплинга рассказывается о жизни человека среди диких зверей. Точнее сказать ребенка, который вырос и сформировался в совершенно не человеческих условиях....
Черный медведь, или барибал, обитает на территории Северной Америки, включая Канаду, США и Мексику. Считается, что сейчас существует 200 000 особей. Их ареалом...
Медведи считаются сравнительно молодым видом, если сопоставлять их с другими разновидностями представителей животного мира. По исследованиям ученых, первые медведи, а вернее их предшественники...
Белый медведь — крупнейший представитель семейства медвежьих. Несмотря на то, что взрослые медведи могут достигать трехметровых размеров и весить почти тонну, назвать их...
Одним из самых опасных и свирепых хищников на планете является медведь гризли. Образ именно этого млекопитающего используется в кинематографе в качестве безжалостного убийцы...
Род медведей насчитывает четыре вида, у трёх из которых (чёрный медведь, белогрудый и бурый медведи) имеются подвиды. Единственный подвид белого медведя, гигантский белый...
Североамериканского Медведя гризли, можно назвать обыкновенным подвидом бурого медведя, который обитает у нас в России. С большой уверенностью можно говорить, что гризли на...
Когда появился наш символ нации? Палеонтологи утверждают, что медведи появились около пяти-шести миллионов лет назад. Сперва это было небольшое животное, обитавшее на территории...
Панду довольно часто величают бамбуковым медведем, хотя, как ни странно, она относится к семейству енотовых. Так же, иногда, панду относят и к семейству...
Благодаря образам медведей, используемых в мультипликациях, у некоторых могло сложиться впечатление, что эти животные безобидны и любят играть. Конечно есть и такие виды:...
Памятники косолапым люди стали устанавливать лишь в ХХ веке. Обычно это монументы, посвященные медведю как символу чего-то; или же зверю, чем-то заслужившему историческую...
Черный медведь или как его принято называть в научном свете — барибал — это вид медведей, который обитает на американском континенте. Черный медведь,...
Огромный хищник, предпочитающий жить на дрейфующих льдах, прекрасно чувствующий себя в воде – так можно описать белого медведя. Несмотря на то, что излюбленной...
На протяжении многих веков панда остается национальным символом и любимым животным в Китае. Большая панда это уникальное млекопитающее, относящееся к семейству медвежьих, обитает...
Медведи – самые крупные из современных хищных зверей. Они могут ходить на задних лапах, быстро бегать и хорошо плавать. У медведей замечательно развито...
Бурый медведь – один из крупнейших современных хищников. По образу питания он наиболее близок к человеку. Всеядность позволяет с успехом содержать животных в...
Царь северной природы, величественный медведь, несмотря на свою силу и выносливость, является редким животным. Помимо повышенного интереса браконьеров к этому виду, ситуации способствует...
Медведи — удивительные животные, несмотря на массу своего тела, они с легкостью карабкаются по деревьям. носятся галопом. обожаемы и детишками, и их родителями....
В опровержение к бытующему мнению, медведи не впадают в анабиоз в зимней спячке. Просто у него замедляются процессы обмена, он начинает реже дышать,...
В России обитает около семи отдельных видов медведей, находящихся в разных частях страны. Чаще всех возможно встретить Бурого и Белого медведя, а также...
Собираясь в отпуск с детьми, родители часто продумывают всю программу развлечений. Некоторые весь отпуск проводят на территории отеля, наслаждаясь морем, а некоторые активно...
С приходом нано-технологий, все чаще можно встретить гуляющих по зоопаркам с новомодными телефонами, планшетами. Некоторые используют технику для звонков или общения в сети-интернет,...
Окуня можно ловить с использованием различных искусственных приманок, которые сегодня представлены в широком ассортименте. Кроме того, в качестве насадки на крючок подойдет обычный...
В настоящее время все более популярной становится процедура протезирования зубов. Современная медицина создала зубные протезы, которые практически невозможно отличить от настоящих, родных зубов....
В последние годы увеличилось количество путешественников по заповедникам и мировым зоопаркам с медведями. Зачастую, чтобы путешествие было насыщенным и комфортным необходимо не только...
Трутовик лакированный или больше известный, как гриб Рейши Ганодема, очень распространен в восточной медицине. Слово «рейша» в японском языке обозначает «духовная сила» и...
В современном мире никого не удивишь различными коммуникациями и развитой инфраструктурой. Вот и в коттеджном поселке Балатон с этим все в полном порядке....
Среднестатистический человек находится в спящем состоянии практически треть жизни, поэтому крайне важно на каком матрасе он спит. Идеальным средством для отдыха является ортопедический...
Каждый из нас, наверняка, попадал в ситуацию, когда только купленная техника внезапно перестает работать. Что делать и как поступить лучше всего, попробуем выяснить....
Многие дамы любят готовить, они с удовольствием радуют своих родных вкусными пирожками и ароматными булочками. Так почему бы любимое занятие не превратить в...
Вопрос об организации укомплектованной жилплощади для своего маленького любимца встает перед каждым владельцем хомячка, кролика или другого представителя отряда грызунов. Домик для питомца...
Геморрой — это острое воспаление прямой кишки. Оно напрямую связанно с инфекцией, которая образуется на стенках кишечника, размножается, там вызывая тем самым внутренний...
Бурый медведь – опасный хищник наших широт. Большинство представителей вида обитают в Азии. Самые крупные особи встречаются на Дальнем Востоке. Большинство людей путают...
Образование в Великобритании по праву считается одним из самых лучших и престижных во всем мире, будь то курсы изучения и совершенствования английского языка,...
Красноярский край известный своим природным заповедником «Столбы». Главной достопримечательностью являются скалы, общее название которых – «столбы». Различают еще и «дикие столбы» – скалы,...
Среди любителей отдыхать в зимний период найдется большое количество желающих получить яркие и новые впечатления от посещения других стран, где есть возможность выбора...
Краснодар – это очень красивый, культурный, исторический и промышленный центр. Сюда постоянно съезжаются туристы с России и соседних стран. Кроме того, город привлекает...
Мода переменчива, как настроение женщины. Вчера она диктовала винтажный стиль, сегодня – классический. Казалось бы, только в моду вошли длинные шарфы, как им...
Упрощенная легализация путём проставления штампа «апостиль» применяется для того, чтобы документы, выданные в своей стране стали действительны на территории другой страны. Этот штамп...
С определенной целью будущий абитуриент делает выбор, в каком университете ему учиться. Главный жизненно нужный и важный шаг каждого человека уверенно стремиться к...
Вопрос верного выбора чемодана стоит перед всеми людьми, особенно перед теми, кто часто путешествует и нуждается в постоянной транспортировке необходимых вещей. Каким же...
Активный отдых зимой включает катание на лыжах, коньках, санках, сноубординг, тюбинг и так далее в зависимости от тог, насколько вы экстремальный человек, от...
Строение зубов у разных зверей имеет много общего, различие зависит от образа жизни и питания. Человек обладает знаниями и может позаботиться о себе...
Изучение иностранного языка довольно непростое занятие и требует много усилий и времени. Китайский язык не считается одним из легких, но все, же если...
Лето, пора не только веселого отдыха, но и больших изматывающих испытаний, под названием «поступление в ВУЗ». Выпускники практически всей страны мечтают поступить в...
Написание диссертации – это, безусловно, самый важный этап при получении кандидатской степени. Однако он не является самым главным, так как самое главное –...
Из самого названия, керамогранит, становится ясно, что это искусственный камень на основе глины. Причем глина используется двух видов: пластичная и не пластичная. Перед...
Обществознание – это наука, изучающая, разнообразные стороны жизни общества, его развитие и место, занимаемое в нем человеком. Обществознание как наука, включает в себя...
Обладатели дачных участков обычно стремятся всеми доступными способами украсить их, а также создать комфортные зоны отдыха. Кто-то идет по пути наименьшего сопротивления –...
Беларусь является одним из самых красивых уголков земли. Невероятно чистый воздух и очаровательные пейзажи, дополненные чистейшими озёрами и красивыми реками, манят туристов посетить...
Каждому из нас приходится частенько проделывать длительные пешие прогулки, и это не обязательно туристические походы, поездки на море, или пикники на природе. Любой...
Стремительно развивающаяся экономика, налоговые платежи, аудит – за всеми этими словами прячется кропотливый труд сотен и тысяч отечественных бухгалтеров. Эта профессия, с одной...
Бытует мнение, что со временем находясь в семье, собака приобретает черты хозяина, равно как привычки хозяина влияют на «собачью жизнь». Одна из таких...
В наше время стремительного развития экономики необходимым становиться получение высшего образования. Многие предпочитают и ставят перед собой цель обучения именно в зарубежных странах....
Те путешественники, которые желают совместить пляжный отдых с насыщенной культурной и развлекательной программой, посетить самый популярный в Европе парк развлечений «Порт Авентура» и...
В XXI веке аллергия распространилась до небывалых масштабов – сегодня практически каждый человек подвержен аллергическим реакциям от того или иного продукта. В первую...
Каждый раз, когда приходит очередной праздник вопрос: «Что подарить?» вводит человека в лёгкий транс, а особенно, если речь идёт о Дне защитника Отечества...
В данный период времени начинаются самые тяжелые дни беременности. Потому что на последних неделях 8-го месяца будущую маму начинают тревожить разнообразные раздражители, так...
В современном мире сложилось так, что английский язык стал основным средством международного общения. По статистике им пользуются больше миллиарда человек на планете, но...
Красивая шкура медведя украсит гостиную или библиотеку, будет служить предметом вашей гордости и вызывать удивление у посетителей. Купить в наше время шкуру медведя...
Парки водных развлечений еще несколько лет назад были доступны нашим соотечественникам только на отдыхе за рубежом. В наши дни комплексы аквапарков существуют в...
Современная жизнь ставит перед человеком необходимость знания иностранного языка. Когда и как должно происходить обучение английскому языку — вопрос, на который нельзя ответить...
В современном обществе образование занимает важную роль в жизни каждого человека. Поэтому каждый человек стремится получить высшее образование для реализации своих планов, карьерного...
Опухоль – это нерегулируемое организмом, патологическое разрастание ткани. Опухоль состоит из непрерывно размножающихся качественно изменившихся клеток животного, передающих свои свойства другим клеткам. А...
Бизнес тренинг очень разнообразен, нельзя выделить какую-то основную область, в которой развивается человек, проходящий его. Основная идея заключается в следующем: научить человека тому,...
Многие владельцы кормят своих любимцев домашней пищей. Для этого им не надо ежедневно ходить в магазин для приобретения корма. Но не всегда домашние...
Каждый человек в своем доме хочет иметь все удобства для приятного времяпровождения. Ежедневно человек потребляет большое количество воды. Поэтому необходимо обустроить дом таким...
Большинство мужчин не хотят отказывать себе в удовольствии постоянно посещая места, где можно поохотиться и половить рыбу. Для такого отдыха необходимо правильно подобрать...
Эта страна славится тем, что количество людей, которые приезжают в это государство, с каждым новым годом только увеличивается. В действительности, эта прекрасная страна,...
Сегодня профессия модельера или дизайнера одежды особенно актуальна, несмотря на то, что далеко не нова. Индустрия моды динамично развивается, а хороших специалистов в...
Петербург, Петроград, Ленинград… А сейчас Санкт-Петербург. Великий город, основанный Петром. Но еще задолго до прихода Петра I, решившего именно здесь прорубить ″окно в...
В Китае традициям чаепития уже несколько тысячелетий. Одной из таких древнейших чайных церемоний является сбор, заготовка, и заваривание чая сорта Пуэр, или как...
Японский язык… Загадочный, непонятный, но очень притягательный и манящий. Популярности его в немалой мере способствовало всеобщее распространение японских анимационных картин. Реально ли человеку,...
Для тех, кто хочет окунуться в райскую, тропическую атмосферу прекрасно подойдет остров Самуи, который считается одним из самых популярных и больших островов Таиланда....
Не так уж и давно в продаже появился очередной телефон фирмы Samsung, продлевающий получившую невероятную популярность линейку смратфонов Galaxy. Это Samsung Galaxy Ace,...
Медвежья жизнь чрезвычайно интересна, материала хватит на составление полноценной научной диссертации. Но даже небольшой реферат на такую тему негоже писать сухо, скупо и...
Знание английского языка сегодня является необходимостью, как знание латыни в Средневековье. Английский стал международным, общепринятым языком на планете. Поэтому все больше людей заинтересованы...
Профессия ветеринара возникла достаточно давно, необходимость в лечении животных появилась еще во времена их первого одомашнивания. Сегодня ветеринары достаточно востребованы на рынке труда,...
Недавно в целях исследования бурых медведей на территории России было применена процедура дистанционной слежки за некоторыми особями с помощью радиосредств. Вместе с традиционными...
Идеей создания зоопарка в Новосибирске, а также первыми шагами по её воплощению был озадачен писатель-зоолог Зверев М.Д. Так в далёком 1933 в городе...
Медведи относятся к крупным млекопитающим, которые обитают по всей планете. Оплодотворение и вынашивание потомства у этой группы животных, как и у всех остальных,...
Созвездие Большой и Малой медведиц знакомо даже людям, далеким от астрономии. На территории России его можно наблюдать невооруженным взглядом круглый год. Но чтобы...
Слова Арктика и медведь – это не просто ассоциации человеческого воображения. Этот материк получил свое название от греческого слова «арктос» (медведь) благодаря созвездию...
На земле нет более крупного хищника, чем медведь. Мощное сложение и рацион питания отчетливо выделяют их среди других представителей хищников. Медведи употребляют в...
Как часто, придя в зоопарк и рассматривая огромных животных наподобие медведей, гиппопотамов, жирафов, мы умиляемся их мощи и внушительным размерам, но редко кто...
Одним из самых распространенных хищных млекопитающих на земле являются медведи. Ареал обитания этих млекопитающих очень широкий. Различные виды медведей можно встретить в Европе,...
Медвежья желчь входила в составы лечебных препаратов на протяжении многих веков. Даже известные врачи древней медицины, такие как Гиппократ, Авиценна, Гален писали о...
В Уссурийском крае и Приамурье обитает удивительный медведь белогрудка – уссурийский медведь. От своего «старшего» сородича бурого медведя его отличает более стройное телосложение,...
Мне, по случаю, пришлось побывать на охотничьей базе, где содержали разных животных для притравки охотничьих собак. Строительные компании потрудились на совесть, база построена...
Исследования, проводимые учеными на Аляске, выявили несколько интересных особенностей продолжительного медвежьего сна. В роли подопытных экземпляров были выбраны пятеро медведей, которых оснастили множеством...
Медведи относятся к самой крупной современной популяции хищных зверей. Длиной до трех метров и массой иногда до тонны эти животные внешне создают весьма...
Все меньше медведей остается в лесах Европы, которые в свое время были основным местом обитания бурых медведей на континенте. Те медведи, которые живут...
Белый медведь (научное название: Ursus maritimus) ведет свои истоки от предков бурого медведя, который стал постепенно обесцвечиваться и приспосабливаться к суровой северной среде...
По величине белогрудый медведь уступает бурому, и телосложение у него стройнее. Вытянутая токая мордочка с большими ушами. У животного блестящий чёрный мех и...
Одним из самых крупных животных нашей полосы является бурый медведь. Отдельные особи достигаю веса в 450 килограммов, но наиболее крупный медведь это камчатский,...
Перед спячкой, медведь подготавливает берлогу к зимнему сну, делает подстилку из травы и мха. По выдранным участкам мха можно примерно определить местонахождение берлоги....
Медведи удивительно любопытные животные, их строение тела чем то напоминает человечье. А вы знали что медведь без шкуры напоминает человеческий облик. Многие охотники...
Многие смотрели мультфильм, назывался он Кунг фу панда, где большая панда, неуклюжая панда, учится боевым искусствам и борется со злом. Хороший и добрый...
Мир для белого медведя ограничен ледяными полями, и этим в первую очередь определяются особенности его поведения. Если судить по животным, содержащимся в неволе,...
За исключением времени, проводимого в берлогах, вся жизнь белого медведя проходит в кочевках. Однако, несмотря на осуществление в последнее время массового мечения зверей,...
В последние десятилетия вопрос о пространственной структуре вида занимает в исследованиях по белому медведю едва ли не центральное место. И это естественно, поскольку...
Год выпуска: 2006 Страна: Великобритания Жанр: Документальный Продолжительность: 00:44:09 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Philip Savoie Эта история разворачивается на Диком...
Год выпуска: 1999 Страна: США Жанр: Документальный Продолжительность: 00:51:49 Перевод: Профессиональный (одноголосый) Описание: Перед вами летопись жизни белой медведицы и двух ее детенышей...
Год выпуска: 1999 Страна: Франция Жанр: документальный Продолжительность: 50мин Режиссер: Жана Масколо де Филлипи. Описание: Джунгли Амазонии — именно здесь, в этих волшебных...
Год выпуска: 2005 Страна: США Жанр: Документальный Продолжительность: 00:39:42 Перевод: Профессиональный (одноголосый) Описание: Медведи и волки: нам кажется, что мы все о них...
Год выпуска: 1994 Страна: США Жанр: Документальный Продолжительность: 52:05 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Вольфгант Байер Описание: В этом фильме Вы...
Год выпуска: 2006 Страна: США, National Geographic Жанр: Научно-популярный, документальный Продолжительность: 00:46:55 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Стивен Тейли Описание: Медведи,...
Пищеварительная система белого медведя исследована недостаточно. В соответствии с характером питания его пищеварительный тракт, в отличие от такового бурого медведя, несет черты большей...
Мускулатура белого медведя практически не исследована (этой теме была посвящена, по-видимому, лишь небольшая специальная работа. При поверхностном знакомстве со зверем обращает на себя...
Год выпуска: 1975 Страна: СССР Жанр: научно-популярный Продолжительность: 54 минуты Перевод: Не требуется Русские субтитры: нет Режиссер: Ю.Ледин Описание: Текст читает: Зиновий Гердт...
Белый медведь, очевидно, может считаться наиболее крупным из ныне живущих наземных хищников: длина тела (без хвоста) взрослых самцов обычно 200— 250 см, редкие...
Белый медведь заметно отличается от бурого удлиненным и более узким, особенно в передней части, туловищем, длинной подвижной шеей, относительно небольшой головой, нешироким лбом,...
Год выпуска: 2008 Страна: Тайвань Жанр: Документальный,познавательный Продолжительность: 00:46:57 Перевод: Профессиональный (одноголосый) Русские субтитры: нет Режиссер: Эндрю Яо / Andrew Yao Описание: Тайваньский...
Год выпуска: 2001 Выпущено: Primesco Productions inc. Жанр: Документальный Продолжительность: 00:39:13 Перевод: Русский любительский одноголосый закадровый перевод Субтитры: нетРежиссер: Дэвид Ликли / David...
Основная причина сокращения численности белого медведя — его промысел, который имеет давнюю историю. Уже в XIV—XV вв. поморы, мало интересуясь китами, специализировались на...
Изучая белых медведей, ученые выяснили, что они предпочитают держаться поодиночке. Это не касается семьи, состоящей из медведицы с ее потомством, у них прекрасно...
Глобальное потепление. Это словосочетание пугает всех жителей планеты Земля, а в частности Исландию. Для местных рыбаков становится все опаснее выходить на рыбалку, поскольку...
Зимой, в некоторых странах мира, 27 февраля отмечается День белого – полярного медведя. Исходя из данных Всемирного Фонда Дикой Природы (WWF), на данный...
Нам этот мишка знаком с детства. Мы знаем его как: мохнатого, косолапого, неуклюжего увальня с непомерной силой в когтистых лапах и крепким здоровьем,...
Что может существовать общего меж медведями и астронавтами? Как оказалось, их повадки увлекательны для тех, кто уже в данный момент начинает думать о...
На днях азиатский (лунный) медведь трагично закончил своё существование… В японском городе Такаяма (префектура Гифу) на автобусной остановке стояла группа людей (которая состояла...
Он имеет три названия: черный медведь, североамериканский черный медведь, барибал. Черный медведь распространен в горных и равнинных лесах, он хорошо приспособился к жизни...
Беременность у самки белого медведя продолжительностью 6 – 7 месяцев (230-250 дней). По истечению этого периода на свет появляются маленькие медвежата. Число детенышей...